Circadian_Clock_in_Mammals
发布时间:2019-12-10 09:44 来源:SABiosciences
- 通路
- 概述
Review
Circadian clocks are molecular time-keeping mechanisms that reside in a diverse range of cell types in a variety of organisms. The primary role of these cell-autonomous clocks is to maintain their own 24-hour molecular rhythm and to drive the rhythmic expression of genes involved in physiology, metabolism and behavior. The ability of the clock to persist in the absence of environmental cues provides internal temporal organization so that rhythmic activities can occur at characteristic times during the circadian cycle. In addition, two other clock properties, entrainment (that is, setting the clock to local time with respect to environmental cycles) and temperature compensation (that is, the ability of the clock to run at the same rate at different temperatures) ensure synchrony with the environment. The importance of the circadian clock is underscored by its ubiquity; clocks are present in organisms ranging from prokaryotic and eukaryotic microbes to plants, insects and mammals (Ref.1). In
Cyanobacteria and Arabidopsis, the circadian oscillator directs transcription of the photosynthetic machinery to the daylight hours, thereby ensuring efficient assimilation of light energy. In mammals, circadian consolidation of locomotor activity to time of food availability and predator avoidance functions to improve survival (Ref.2).
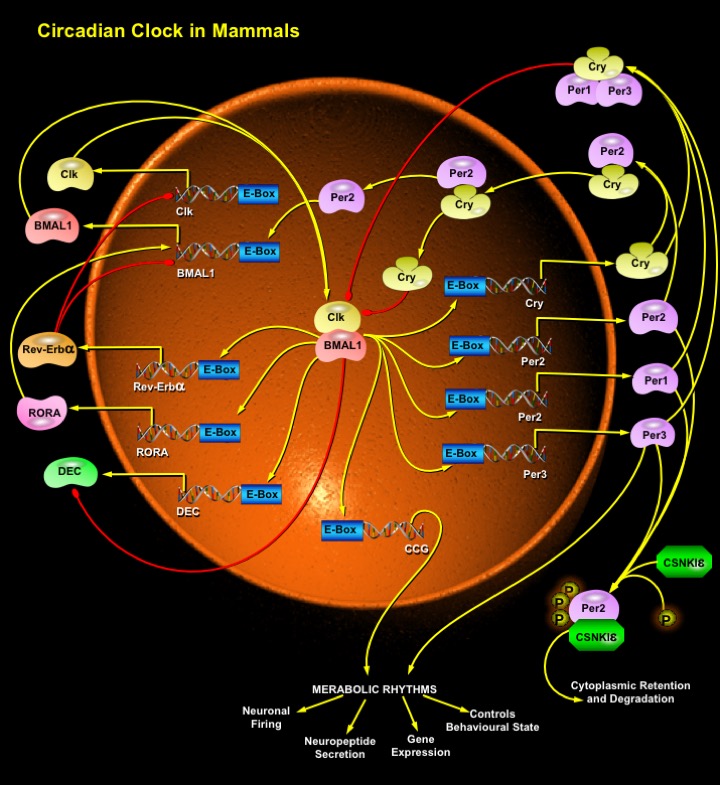
The mammalian circadian system is critical for the proper regulation of behavioral and physiological rhythms. The central oscillator, or master clock, is located in the hypothalamic SCN (Suprachiasmatic Nucleus) (Ref.3). Additional circadian clocks are dispersed throughout most organs and tissues of an animal. The most prominent stimuli capable of synchronizing circadian oscillations to the environment is light. This occurs through daily photic signaling to the SCN, which ultimately results in the appropriate phasing of the various biological rhythms. The circadian clock has been conceptualized as a series of three components: an 'Entrainment Pathway' that transmits environmental (usually light) signals to the timekeeping apparatus; a timekeeping apparatus, or 'Oscillator', which operates in the absence of environmental cues and is the core component of the circadian clock; and 'Output Pathways' that are activated at specific times of the Circadian Cycle by the Circadian Oscillator (Ref.2). In mammals, five structures play important roles in the circadian regulation of physiology and behavior. These structures are the photosensitive retina, the RHT (Retinohypothalamic Tract), the SCN, the pineal gland, and the peripheral oscillators. In mammals the SCN functions as the central or master clock. In addition to receiving photic input signals from the retina, the hypothalamic SCN also receives feedback from targets such as the pineal gland. Circadian signals that originate from the SCN can act directly or indirectly to maintain proper oscillations in a variety of clock targets (Ref.1). The SCN are synchronized to the 24hr day by the daily light-dark cycle and in turn synchronize rhythms in behavior (e.g. locomotor activity), by secreting factors (e.g. TGF-Alpha and Prokineticin) that act locally within the hypothalamus, and entrains subservient circadian oscillators in peripheral tissues (e.g. liver and kidney) via humoral signals. Such peripheral oscillators can, however, become uncoupled from the SCN if their specific needs dictate—as occurs in liver, lung and skeletal muscle after entrainment by food (Ref.5). The positive transcriptional elements in the circadian oscillator include two bHLH (basic Helix-Loop-Helix) PAS-domain containing transcription factors, Clk (Clock) and BMAL1 (Brain and Muscle ARNT-Like-1). In the SCN neurons, the intracellular levels of Clk remain steady throughout the 24-hour period, whereas BMAL1 expression levels are high at the beginning of a subjective day and low at the beginning of a subjective night. The high level of BMAL1 promotes the formation of BMAL1–Clk heterodimers. These heterodimers drive the transcription of three mammalian period genes Per1, Per2 and Per3, two Cryptochrome genes Cry1 and Cry2 and Rev–Erb-Alpha genes by binding to their respective CACGTG E-Box elements present in the promoters of other Central Clock Genes and CCG (Clock-Controlled Genes). The BMAL1–Clk heterodimer can also inhibit BMAL1 transcription. After transcription and translation, the Rev–Erb-Alpha protein enters the nucleus to suppress the transcription of BMAL1 and Cry genes. A second cluster of genes are driven by RORA (RAR-Related Orphan Receptor-A), which are sensitive to negative regulation by Rev–Erb-Alpha and so are expressed in phase with BMAL1 (Ref.6). Rev-Erb-Alpha/RORA is involved in gene expression during circadian night, which is in phase with BMAL1 and in antiphase to Per2 oscillations. As the Per proteins, such as Per2, accumulate in the cytoplasm, they become phosphorylated by CSNK-I-Epsilon (Casein Kinase-I-Epsilon). The phosphorylated forms of Per are unstable and are degraded by ubiquitylation. Late in the subjective day, however, Cry accumulates in the cytoplasm, promoting the formation of stable CSNK-I-Epsilon/Per/Cry Complexes, which enter the nucleus at the beginning of a subjective night. Once in the nucleus, Cry1 disrupts the Clk/BMAL1-associated transcriptional complex, resulting in the inhibition of Cry, Per and Rev–Erb-Alpha transcription, and derepression of BMAL1 transcription. The interacting positive and negative feedback loops of circadian genes ensure low levels of Per and Cry, and a high level of BMAL1 at the beginning of a new circadian day.
Recently, repressive transcription factors, DECs (Differentially Expressed in Chondrocytes), have been shown to repress Clk/BMAL1-induced activation of the Per1 promoter through two distinct mechanisms: competing for E-Box binding and interacting with BMAL1. DEC proteins belong to a new and structurally distinct class of bHLH transcription factors that interact with Class-B E-Box (Ref.7).
In the forebrain the function of Clk is replaced by NPAS2 (Neuronal PAS domain protein-2, also called MOP4) (Ref.8). In addition, a number of posttranslational events, such as the control of protein phosphorylation, degradation, and nuclear entry, contribute critically to the generation of daily oscillations in clock gene products (Ref.9). As circadian pacemakers can measure time only approximately, their phase has to be adjusted every day in order to stay in synchrony with geophysical time. Many problems in sleep patterns are related to an inability to properly entrain the circadian clock to the day-night cycle. Furthermore, disruptions of the sleep pattern due to jet lag or shift work are also related to the entrainment of the circadian clock (Ref.10). Abnormality in circadian behavior may cause FASPS (Familial Advanced Sleep Phase Syndrome). People with FASPS are referred to as “morning larks” – they sleep for a normal amount of time, but sleep onset occurs around 7:30 p.m., and they awaken around 4:30 a.m. FASPS is caused by a mutation in the CSNK-I-Epsilon binding region of Per2 that leads to decreased phosphorylation of Per2 by CSNK-I-Epsilon (Ref.11, 12, 13).
References
- 1
- Panda S, Antoch MP, Miller BH, Su AI, Schook AB, Straume M, Schultz PG, Kay SA, Takahashi JS, Hogenesch JB. Coordinated transcription of key pathways in the mouse by the circadian clock.
- 2
- Nitabach MN. Circadian rhythms: clock coordination.
- 3
- Aton SJ, Herzog ED. Come together, right...now: synchronization of rhythms in a mammalian circadian clock.
- 4
- Moore RY. A clock for the ages.
- 5
- Yamazaki S, Numano R, Abe M, Hida A, Takahashi R, Ueda M, Block GD, Sakaki Y, Menaker M, Tei H. Resetting central and peripheral circadian oscillators in transgenic rats.
- 6
- Guillaumond F, Dardente H, Giguere V, Cermakian N. Differential control of Bmal1 circadian transcription by REV-ERB and ROR nuclear receptors.
- 7
- Li Y, Xie M, Song X, Gragen S, Sachdeva K, Wan Y, Yan B. DEC1 negatively regulates the expression of DEC2 through binding to the E-box in the proximal promoter.
- 8
- Lee C, Etchegaray JP, Cagampang FR, Loudon AS, Reppert SM. Posttranslational mechanisms regulate the mammalian circadian clock.
- 9
- Field MD, Maywood ES, O\'Brien JA, Weaver DR, Reppert SM, Hastings MH. Analysis of clock proteins in mouse SCN demonstrates phylogenetic divergence of the circadian clockwork and resetting mechanisms.
- 10
- Schibler U, Sassone-Corsi P. A web of circadian pacemakers.
 关于我们
关于我们