TCR_Signaling
发布时间:2019-12-11 14:48 来源:SABiosciences
- 通路
- 概述
Review
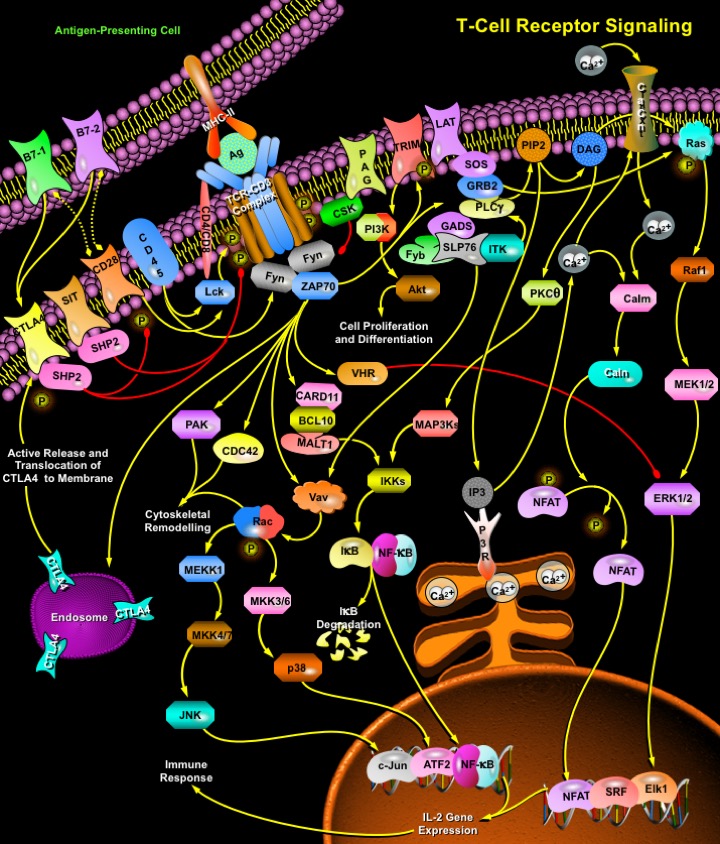
T-Cells are a subset of lymphocytes that play a large role in the immune response. The TCR (T-Cell Receptor) is a complex of integral membrane proteins that participates in the activation of T-Cells in response to the presentation of antigen. Stimulation of TCR is triggered by MHC (Major Histocompatibility Complex) molecules on Antigen Presenting Cells that present antigen peptides to TCR complexes and induce a series of intracellular signaling cascades. Engagement of the TCR initiates positive (signal-enhancing) and negative (signal-attenuating) cascades that ultimately result in cellular proliferation, differentiation, cytokine production, and/or activation-induced cell death. These signaling cascades regulate T-Cell development, homeostasis, activation, acquisition of effector’s functions and apoptosis (Ref.1 & 2).
TCR is composed of six different chains that form the TCR heterodimer responsible for ligand recognition. CD3 molecules (CD3-Gamma, Delta, Epsilon and Zeta), which are assembled together with the TCR heterodimer, possess a characteristic sequence motif for tyrosine phosphorylation, known as ITAMs (Immunoreceptor Tyrosine-based Activation Motifs). The TCR polypeptides themselves have very short cytoplasmic tails, and all proximal signaling events are mediated through the CD3 molecules. T-Cell activation is initiated by the interaction of the TCR with antigenic peptides complexed to MHC-II molecules, and accessory proteins essential for MHC-II. TCR-CD3 complex interaction plays an important role in mediating cell recognition events. TCR engagement by antigens triggers the tyrosine phosphorylation of the ITAMs, present in the TCR-associated CD3-Zeta subunits. Such ITAMs function by orchestrating the sequential activation of the Src-related PTKs: Lck and Fyn, which initiate TCR signaling, followed by that of ZAP70, which further amplifies the response. Lck is activated by the interaction of MHC-II and CD4 or CD8. These various PTKs induce tyrosine phosphorylation of several polypeptides, including the transmembrane adaptors: LAT (Linker Activator for T-Cells) and TRIM (T-Cell Receptor Interacting Molecule). Protein tyrosine phosphorylation subsequently leads to the activation of multiple pathways, including ERK (Extracellular Signal Regulated Kinase), JNK (c-Jun N-terminal Kinase), NF-KappaB (Nuclear Factor-KappaB) and NFAT (Nuclear Factor of Activated T-Cells) pathways, which ultimately induce effector functions (Ref.3, 4 & 5).
TCR activation is regulated by various costimulatory receptors. CD28 provides an essential co-stimulatory signal during T-cell activation, which augments the production of IL-2 (Interleukin-2), increases T-Cell proliferation and prevents the induction of anergy and cell death. Once ligated by B7-1 or B7-2, CD28 provides the T-Cell with an initial adhesion capable of approximating the T-Cell and Antigen Presenting Cell membranes. Besides CD28, many other transmembrane receptors also modulate specific elements of TCR signaling. CD45 (CD45 Antigen) is one such receptor which regulates TCR signaling by modulating the phosphorylation state of the tyrosine kinases like Lck and Fyn, and antagonizing the inhibitory impact of inhibitory proteins, thereby favoring T-Cell activation. Lck is also activated by CD28. Activated Lck in turn activates the phosphorylation and activation of the TCR-CD3 complex and consequently, the tyrosine kinases: Fyn and ZAP70 (Zeta-Chain (TCR) Associated Protein Kinase of 70 kDa) (Ref.3). ZAP70 remains associated with the TCR-Zeta chain and induces activation of LAT (Linker for Activation of T-Cells) and TRIM. TRIM contributes to TCR-dependent PI3K-->Akt signaling, culminating in cell survival and differentiation. LAT is an integral membrane adaptor protein that resides in lipid membrane rafts and binds to the adaptor GADS (Growth Factor Receptor-Bound Protein-2-Related Adaptor Protein-2), SLP76 (SH2 Domain-Containing Leukocyte Protein-76), ITK (IL-2 inducible T-cell kinase), and Fyb. LAT stimulates one critical protein PLC-Gamma1 (Phospholipase-C-Gamma1) that is responsible for the production of the second messengers DAG (Diacylglycerol) and IP3 (Inositol Triphosphate) by cleaving PIP2 (Phosphatidylinositol-4, 5-Bisphosphate) at the plasma membrane. DAG activates a number of proteins, such as PKC-Theta (Protein Kinase-C-Theta) and Ras, whereas IP3 binds to IP3R (IP3 Receptor) on the surface of the ER (Endoplasmic Reticulum) and releases Ca2+. This event triggers the opening of CRAC (Ca2+ Release Activated Ca2+ Channels) at the plasma membrane, allowing influx of extracellular Ca2+. The increased Ca2+ levels then activate the protein phosphatase Calcineurin by disrupting the inhibitory effects of Calm (Calmodulin). Calcineurin activation leads to the dephosphorylation of NFAT, allowing it to enter the nucleus, where it cooperates with other transcription factors to bind promoters (Ref.1, 2, 6 & 7).
In addition to enzymes, tyrosine-phosphorylated LAT also binds multiple members of the GRB2 family of adaptor proteins, such as GRB2, GRAP (GRB2- Related Adaptor Protein) and GADS (GRB2 -Related Adaptor Protein-2) to facilitate the assembly of macromolecular signaling complexes that are required for efficient T-Cell activation. The interaction of tyrosine-phosphorylated LAT with GRB2 provides a mechanism by which GRB2 and GRAP-associated SOS are recruited to the plasma membrane and potentially activate Ras (Ref.6). Activation of Ras leads to the activation of a number of serine/threonine kinases: Raf1, MEK (MAPK/ERK Kinase) and dual-specificity kinases that are responsible for the eventual activation of the MAPKs (Mitogen Activated Protein Kinases): ERK1/2. In contrast, ERK activation can also be downregulated by TCR signaling by phophorylated VHR (Dual specificity protein phosphatase VHR), a dual specific phosphatase, phosphorylated by ZAP70. Other MAPKs like JNK (c-Jun N-terminal Kinase) and p38 are also activated by ZAP70 and SLP76, downstream of Vav. Vav in turn phosphorylates and activates the GTP-binding protein Rac which furher mediates the activation of MEKK1 (MAP/ERK Kinase Kinase-1). p38 is activated by MEKK1 through the activation of MKK3 (Mitogen-Activated Protein Kinase Kinase-3)/MKK6. JNK is activated by MEKK1 through activation of MKK4 (Mitogen-Activated Protein Kinase Kinase-4)/MKK7. These MAPKs directly phosphorylate transcription factors involved in the formation of the heterodimeric transcription factor, Jun-Fos complex, as well as NFAT and IL-2 (Interleukin-2) gene expression in synergy with Ca2+ signaling. ERKs, JNK and p38 directly phosphorylate the transcription factors: Elk1, c-Jun and ATF2 (Activating Transcription Factor-2) respectively. Another transcription factor important for the generation of IL-2 is NF-KappaB (Ref.6, 7 & 8).
NF-KappaB members control the expression of various genes involved in inflammatory, apoptotic and immune responses (Ref.3). Activation of NF-KappaB is dependent on stimulation of the TCR and co-stimulation via CD28. CARD11, a MAGUK (Membrane-Associated Guanylate Kinase Homolog) family member, downstream of ZAP70 and the MAP3K (Mitogen-Activated Protein Kinase Kinase Kinase), a downstream product of PKC-Theta activation, participate in the activation of the heterotrimeric IKK (I-KappaB Kinases) complex. CARD11 acts in concert with the MALT-lymphoma-associated proteins: BCL10 (B-Cell CLL/lymphoma-10) and MALT1 (Mucosa Associated Lymphoid Tissue Lymphoma Translocation Gene-1), which regulate NF-KappaB via the IKK complex. The IKK complex regulates NF-KappaB activity by phosphorylating I-KappaB (Inhibitor of Kappa Light Chain Gene Enhancer in B-Cells), which leads to its ubiquitination. Free from its association with I-KappaB, NF-KappaB can move into the nucleus and activate transcription. TCR activation also leads to cytoskeletal rearrangements through the activation of GTP-binding proteins Rac, PAK and CDC42, downstream of ZAP70 (Ref.9, 10 & 11) .
Negative regulation of TCR signaling is also of significance, in order to keep a check on the hyperactivation of immune response associated with the pathway. This is achieved through the inthervention of several proteins and receptors as well. SIT (SHP2-Interacting Transmembrane Adaptor Protein) is a recently identified transmembrane adaptor protein, which inducibly interacts with the SHP2 (SH2-containing Protein tyrosine Phosphatase-2) via an ITIM (Immunoreceptor Tyrosine-based Inhibition Motif), and the complex acts as a critical negative regulator of TCR-mediated signaling (Ref.12 & 13). In resting human T-Cells, PAG (Phosphoprotein Associated with Glycosphingolipid Microdomains), a transmembrane adaptor molecule found in lipid rafts, is tyrosine phosphorylated and associated with CSK (c-Src Tyrosine Kinase), an inhibitor of Src-related protein tyrosine kinases. As a result, overexpression of PAG inhibits TCR-mediated responses. These modifications (association of PAG and CSK) are rapidly lost in response to TCR stimulation in order to maintain uninterrupted TCR signaling (Ref.7). CTLA4 (Cytotoxic T-Lymphocyte Antigen-4) negatively regulates T-Cell activation. The transmembrane protein CTLA4 also serves as a natural inhibitor - that once T-Cells become activated by whatever disease process is turning them on, the body has a natural process to turn down the T-Cell pathways so that it does not get too much out of control. After T-Cell activation, CTLA4 is rapidly endocytosed, thus removing it rapidly from the cell surface. The significance of the apparent tight control of CTLA4 expression is due to the fact that CTLA4 has a greater affinity for its B7-1/B7-2 ligands in comparison to CD28; thus, when CTLA4 is not required, it is endocytosed, to maintain a fast T-cell activation response. When a need to control TCR signaling arises, ZAP70 activates the process of active release and translocation of CTLA4 to the membrane. At the membrane, CTLA4 interacts with the SHP2 and inhibits the phosphorylation of TCR. Another mechanism by which CTLA4 might antagonize T-Cell function is through inhibition of CD28 signaling by competing for their shared ligands B7-1 and B7-2 (Ref.14 & 15).
Every effective immune response involves T-Cell activation; however, T-Cells are especially important in cell-mediated immunity, which is the defense against tumor cells and pathogenic organisms inside body cells. They are also involved in rejection reactions. TCR stimulation may provoke different cell responses (proliferation, anergy to subsequent stimuli, cell death) in mature circulating T-Cells, as in thymocytes. Deregulation of T-Cell function, whether by defect or by excess, results in dire consequences for the organism i.e., immunodeficiency and autoimmunity respectively. It is also an extremely sensitive system, as very few peptide-MHC complexes on an antigen-presenting cell are needed to trigger a T-Cell response. Since T-Cells play various critical roles in orchestrating the immune responses, this knowledge should lead to an understanding of how breakdowns in immune regulation lead to autoimmune diseases and of how the immune system could be better manipulated to overcome afflictions such as cancer, infection and autoimmune diseases (Ref.16 & 17).
References
- 1
- Lin J, Weiss A. T cell receptor signaling.
- 2
- Okkenhaug K, Bilancio A, Emery JL, Vanhaesebroeck B. Phosphoinositide 3-kinase in T cell activation and survival.
- 3
- Davis MM. A new trigger for T cells.
- 4
- Coombs D, Kalergis AM, Nathenson SG, Wofsy C, Goldstein B. Activated TCRs remain marked for internalization after dissociation from pMHC.
- 5
- Okkenhaug K, Vanhaesebroeck B. PI3K in lymphocyte development, differentiation and activation.
- 6
- Gong Q, Cheng AM, Akk AM, Alberola-Ila J, Gong G, Pawson T, Chan AC. Disruption of T cell signaling networks and development by Grb2 haploid insufficiency.
- 7
- Davidson D, Bakinowski M, Thomas ML, Horejsi V, Veillette A. Phosphorylation-dependent regulation of T-cell activation by PAG/Cbp, a lipid raft-associated transmembrane adaptor.
- 8
- Alonso A, Rahmouni S, Williams S, van Stipdonk M, Jaroszewski L, Godzik A, Abraham RT, Schoenberger SP, Mustelin T. Tyrosine phosphorylation of VHR phosphatase by ZAP-70.
- 9
- Pomerantz JL, Denny EM, Baltimore D. CARD11 mediates factor-specific activation of NF-kappaB by the T cell receptor complex.
- 10
- Hara H, Wada T, Bakal C, Kozieradzki I, Suzuki S, Suzuki N, Nghiem M, Griffiths EK, Krawczyk C, Bauer B, D\'Acquisto F, Ghosh S, Yeh WC, Baier G, Rottapel R, Penninger JM. The MAGUK family protein CARD11 is essential for lymphocyte activation.
 关于我们
关于我们