Rac1_Pathway
发布时间:2019-12-11 11:28 来源:SABiosciences
- 通路
- 概述
Review
To achieve strong adhesion to their neighbors and sustain stress and tension, epithelial cells develop many different specialized adhesive structures. Breakdown of these structures occurs during tumor progression with the development of a fibroblastic morphology characteristic of metastatic cells. Adhesion receptors of the Cadherin family have been implicated in these cellular processes, which play an important role in the development and maintenance of the differentiated epithelial phenotype during organogenesis and adult life. Cadherin-mediated adhesion requires the activity of the cytosolic proteins of the Rho subfamily members, Rho, Rac and CDC42 (Cell Division Cycle-42). They belong to the Ras Superfamily of small GTPases, whose function is regulated depending on the type of guanine nucleotide bound. Mammals have three closely related Rac isoforms, Rac1, Rac2 and Rac3 (Ref.1).
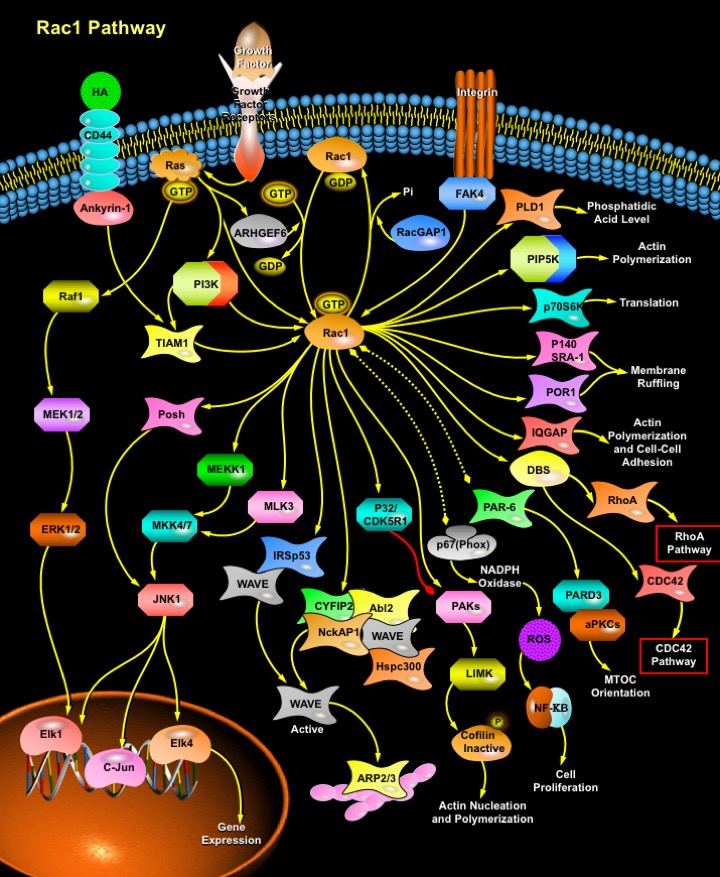
Rac1 is a small G-protein in the Rho family that drives Actin polymerization and formation of lamellipodia, promotes cell-cell adhesion, breakdown and migration of different carcinoma cells. In other epithelial cell lines, Rac activation also plays a role in scattering after distinct stimuli such as Growth factor stimulation or Integrin engagement. It also plays a critical role in processes, such as control of cell morphology, transcriptional activation, and apoptosis signaling. The broad range of events controlled by this GTPase requires regulation of its interactions with multiple downstream targets. Rac1 is activated by GEF (Guanine Nucleotide Exchange Factors), in particular ARHGEF6 and repressed by GAPs (GTPase-Activating Proteins), specifically RacGAP. Rac1 GTPase mediates key cellular processes in response to upstream regulators such as Growth Factors, Integrins and HA (Hyaluronic Acid)-binding receptor CD44. Growth Factors and Growth Factor Receptors stimulate Ras by recruiting SOS (Son of Sevenless), GRB2 (Growth Factor Receptor-Bound Protein-2) and SHC ({SHC (Src Homology-2 Domain Containing) Transforming Protein}) to the membrane in order to activate PI3K (Phosphatidylinositde-3 Kinase) or phosphorylate Src to recruit PI3K. Rac is a key downstream target/effector of PI3K. Rac1 can also be activated by Integrin via FAK (Focal Adhesion Kinase). Interaction between CD44 (the HA-binding Receptor) and TIAM1 (T-lymphoma invasion and metastasis 1) can also activate Rac1. TIAM1 is one of the known GDP/GTP exchange factors for Rac1. TIAM1 and the cytoskeletal protein, Ankyrin, physically associate as a complex in vivo. In particular, the ARD (Ankyrin Repeat Domain) of Ankyrin is responsible for TIAM1 binding. Ankyrin binding to TIAM1 activates Rac1. Upon activation, Rac1 interacts with and regulate a spectrum of functionally diverse downstream effectors, initiating a network of cytoplasmic and nuclear signaling cascades (Ref.2, 3 & 4).
A number of proteins act as targets for Rac1 including PAKs (p21-Activated Kinases), IQGAP1 (IQ motif containing GTPase Activating Protein-1), MRCK/ CDC42BPA (CDC42 binding protein kinase Alpha (DMPK-like), POR1 (Partner Of Rac1) and POSH (Plenty Of SH3s). Rac binds p67(Phox) to increase activation of the NADPH Oxidase system and production of ROS (Reactive Oxygen Species), which mediate activation of NF-KappaB (Nuclear Factor-KappaB)-dependent gene expression, effects of Rac on cell cycle progression and inhibition of Rho activity. Rac binds the WAVE (WASP Family Verprolin Homology Domain-Containing Protein) complex (also containing Abi and IRSp53/58), to release active WAVE, which promotes actin polymerization in lamellipodia through activation of the ARP2/3 (Actin-Related Protein-2/3) complex. Rac also binds to the actin-binding protein IQGAP, which is implicated in regulation of cell-cell adhesion and Microtubule orientation. By binding to the microtubule tip protein Clip170, IQGAP1 captures growing microtubules at the leading edge of migrating fibroblasts, which results in cell polarization. Recently, a novel Rac-interacting protein, POR1 (Partner of Rac1), has been shown to play a role in membrane ruffling. p140SRA1 (Specifically Rac1-Associated Protein) is also a novel Specific target for Rac1 Small GTPase and is also involved in membrane ruffling. Recent findings Rac, involved in the dynamics of Actin cytoskeleton and cell polarity, bind to a protein complex containing PAR-6, PAR-3/ASIP, and aPKC (atypical Protein Kinase-C) (Ref.1, 5 & 6).
Members of the Rho subfamily of GTP-binding proteins including Rho and Rac are also implicated in the regulation of PLD (Phospholipase-D). PLD catalyzes the hydrolysis of Phosphatidylcholine to yield Phosphatidic Acid and Choline. Phosphatidic Acid is a second messenger involved in membrane remodeling events that are critical to cell growth, such as vesicle trafficking and regulated secretion. Both Rac and Rho bind to and activate PIP5K (Phosphatidylinositol-4-Phosphate 5-Kinase), which increases the amount of PIP2 (Phosphatidylinositol (4,5)-bisphosphate). Rac1 coordinately activates p70S6K (p70 Ribosomal -S6 Kinase) and JNK (c-Jun Kinase) via MLK3 (Mixed-Lineage Protein Kinase-3) activation. MLK3 activates JNK via MKK4/7 (MAP Kinase Kinase-4/7). JNK, once activated enter the nucleus and phosphorylate transcription factors such as c-Jun, c-Fos, Elk1 and Elk4. Rac also activates DBS (Dbl's Big Sister), which further activates RhoA and CDC42. In neurons, Rac acts through the protein kinase CDK5 (Cyclin-Dependent Kinase-5) and p35 to phosphorylate and downregulate PAK1, increasing neuronal migration. PAK1 also phosphorylates and activates LIMK (LIM Kinase), which phosphorylates Cofilin as one target. Cofilin stimulates actin depolymerization and changes in cell structure, and phosphorylation of Cofilin by LIMK represses its activity (Ref.7, 8 & 9).
Signaling pathways that are regulated by Rho family members play an important role in several pathological conditions, including cancer, inflammation, and bacterial infections. Rac controls the generation of ROS , both in leukocytes and nonhematopoietic cells and is necessary for Cadherin-dependent adhesion. Rac activation is required for the full transformed phenotype induced by oncogenes such as TIAM1 and Ras. In addition, Rac activation perturbs cadherin contacts with a concomitant change in cell shape, including formation of lamellae/protusions and conversion to a fibroblastic morphology. Translocation of Rac to the plasma membrane is also required for assembly and activation of the NADPH Oxidase complex (Ref.2 & 10).
References
- 1
- Kimura K, Kawamoto K, Teranishi S, Nishida T. Role of Rac1 in fibronectin-induced adhesion and motility of human corneal epithelial cells.
- 2
- Wheeler AP, Wells CM, Smith SD, Vega FM, Henderson RB, Tybulewicz VL, Ridley AJ. Rac1 and Rac2 regulate macrophage morphology but are not essential for migration.
- 3
- Murai T, Miyazaki Y, Nishinakamura H, Sugahara KN, Miyauchi T, Sako Y, Yanagida T, Miyasaka M. Engagement of CD44 promotes Rac activation and CD44 cleavage during tumor cell migration.
- 4
- Adithi M, Venkatesan N, Kandalam M, Biswas J, Krishnakumar S. Expressions of Rac1, Tiam1 and Cdc42 in retinoblastoma.
- 5
- Soderling SH, Scott JD. WAVE signalling: from biochemistry to biology.
- 6
- Natale DR, Watson AJ. Rac-1 and IQGAP are potential regulators of E-cadherin-catenin interactions during murine preimplantation development.
- 7
- Santy LC, Casanova JE. Activation of ARF6 by ARNO stimulates epithelial cell migration through downstream activation of both Rac1 and phospholipase D.
- 8
- Wennerberg K, Ellerbroek SM, Liu RY, Karnoub AE, Burridge K, Der CJ. RhoG signals in parallel with Rac1 and Cdc42.
- 9
- Kesavapany S, Amin N, Zheng YL, Nijhara R, Jaffe H, Sihag R, Gutkind JS, Takahashi S, Kulkarni A, Grant P, Pant HC. p35/cyclin-dependent kinase 5 phosphorylation of ras guanine nucleotide releasing factor 2 (RasGRF2) mediates Rac-dependent Extrac
- 10
- Chrostek A, Wu X, Quondamatteo F, Hu R, Sanecka A, Niemann C, Langbein L, Haase I, Brakebusch C. Rac1 is crucial for hair follicle integrity but is not essential for maintenance of the epidermis.
 关于我们
关于我们