PI3K_Signaling_in_B-Lymphocyte
发布时间:2019-12-11 11:11 来源:SABiosciences
- 通路
- 概述
Review
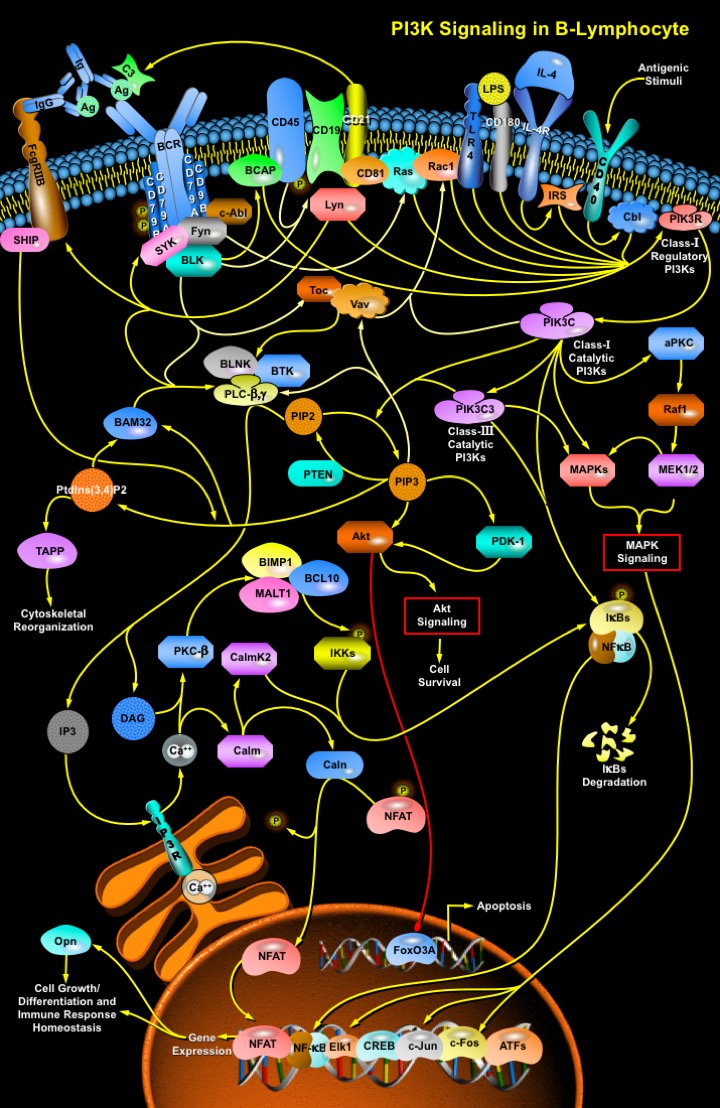
PI3Ks (Phosphoinositide-3-Kinases) regulate numerous biological processes, including cell growth, differentiation, survival, proliferation, migration and metabolism. In the immune system, impaired PI3K signaling leads to immunodeficiency, whereas unrestrained PI3K signaling contributes to autoimmunity and Leukemia. The Class I and III PI3Ks basically facilitate B-cell development through defined stages, resulting in at least three distinct lineages of mature B-lymphocytes. In B-cells, PI3K is activated within seconds of antigen-receptor triggering. The BCR (B-Cell antigen Receptor) plays a critical role in recognition of antigens and activation of B-cells. The BCR or mIg (Membrane Immunoglobulin) is associated with Ig-Alpha/CD79A (CD79A Antigen) and Ig-Beta/CD79B (CD79B Antigen) heterodimer. The mIg subunits bind antigen and cause receptor aggregation, while the Ig-Alpha/Ig-Beta subunits transduce signals to the cell interior (Ref.1 & 2). Engagement of BCR-antigen complex activates intracellular protein tyrosine kinases like SYK (Spleen Tyrosine Kinase), BTK (Bruton Agammaglobulinemia Tyrosine Kinase) and Fyn (Fyn Oncogene Related to Src, FGR, YES). These tyrosine kinases phosphorylate the co-receptors CD19 (CD19 Antigen) and BCAP (B-Cell Adaptor Protein) at the YXXM motifs, which provide binding sites for PI3Ks. CD19 is one of the main regulators of PI3K activity in B-cells. CD19 has an important, but not indispensable, role in PI3K activation rather it is required for sustained PI3K activation after BCR stimulation. The co-receptor complex is also composed of CD21 (CD21 Antigen) and CD81. CD21 binds opsinized antigenic particles and activate Complement Component-C3, a reaction central to complement function in the immune response and sustained BCR signaling. Many other transmembrane receptors are known to modulate specific elements of BCR signaling. A few of these are CD45 (CD45 Antigen) and Fc-GammaRIIB (Low Affinity Immunoglobulin Gamma Fc Region Receptor-IIB). CD45 occurs as a component of a complex of proteins associated with the antigen receptor, and may regulate signal transduction by modulating the phosphorylation state of the co-receptors like CD19 (Ref.2 & 3).
Moreover for B-cell development, the capacity of CD19 to promote thymus-dependent immune responses is linked to its capacity to recruit and activate PI3K. CD19 phosphorylation activates Lyn (V-YES-1 Yamaguchi Sarcoma Viral Related Oncogene Homolog) that in turn recruits regulatory PI3Ks. Lyn activation stimulates BCR and associated protein tyrosine kinases to activate Vav (Oncogene Vav) and Tec (Tec Protein Tyrosine Kinase). Vav and Tec contribute to PI3K activation in B-cells by a mechanism that involves the activation of Rac1 (Ras-Related C3 Botulinum Toxin Substrate-1), which then binds to regulatory PI3Ks through its RhoGAP (Rho-GTPase-Activating Protein) domain. This mechanism places Vav and Tec upstream of PI3K signaling. B-cell proliferation is also stimulated by IL-4 (Interleukin-4)/IL-4R (Interleukin-4 Receptor) through IRS (Insulin Receptor Substrate) activation; LPS (Lipopolysaccharide) stimulated TLR4 (Toll-Like Receptor-4)/CD180 (CD180 Antigen) activation; or a CD40 (B-Cell-Associated Molecule CD40) activated Cbl (Cas-Br-M (Murine) Ecotropic Retroviral Transforming Sequence) that engages p85Alpha-associated p110Delta, thus enhancing PI3K signaling. The regulatory Class I PI3K subunits then activate subsequent catalytic Class I subunits (Ref.4 & 5). Once active the catalytic Class I PI3Ks phosphorylate PIP2 (Phosphatidylinositol 4,5-Bisphosphate) to produce PIP3 (Phosphatidylinositol-3,4,5-trisphosphate) at the inner leaflet of the plasma membrane and also activate the Class III PIK3C3 (Phosphoinositide-3-Kinase-Class-3) to enhance the phosphorylation process. PI3K/PIP3 signaling then turns on Akt Signaling via Akt (v-Akt Murine Thymoma Viral Oncogene Homolog)/PDK-1 (Phosphoinositide-Dependent Kinase-1) activation and down regulation of apoptosis regulating Forkhead family of transcription factors like FoxO3A (Forkhead Box-O3A), and thereby facilitates cell survival. However, the conversion of PIP2 to PIP3 is reversed by PTEN (Phosphatase and Tensin Homolog), a lipid phosphatase, which removes the D3 phosphate from PIP3 (Ref.6). A similar inhibitory function is also exhibited by the Fc-Gamma-RIIB induced SHIP (SH2-Containing Inositol Phosphatase). Upon BCR co-ligation Lyn and associated co-receptors generate an SH2 recognition domain that provides the binding site for the inhibitory signaling molecule SHIP, which removes excess accumulation of PIP3 by converting it to PtdIns(3,4)P2 (Phosphatidylinositol 3,4-Bisphosphate). This leads to the abrogation of PI3K/PIP3 signaling. Thus the PI3K signal is taken over by TAPP (Tandem PH-Domain-Containing Protein) adaptor proteins, which have binding specificity for PtdIns(3,4)P2 and influence the process of cytoskeletal reorganization (Ref.7 & 8).
The conversion of PIP3 to PtdIns(3,4)P2 also terminates activation of Vav, Tec and the Signalosome complex that comprises of BTK (Bruton Agammaglobulinemia Tyrosine Kinase), PLC-Beta (Phospholipase-C-Beta), PLC-Gamma (Phospholipase-C-Gamma) and BLNK (B-Cell Linker Protein). Generation of PIP3 and PtdIns(3,4)P2 activates BAM32 (B-Cell Adaptor Molecule-32KD), an adaptor protein with a high affinity PIP3-binding PH domain that generates BCR-dependent Calcium flux. After BCR ligation, BAM32 is recruited to the plasma membrane through its PH domain. Moreover, BCR-stimulated calcium flux also depends on the Signalosome activation by BCR-associated co-receptors, BAM32, Vav and Tec. Sequential phosphorylation by Lyn and autophosphorylation of BTK contributes to the BCR-stimulated and PI3K-dependent calcium signaling, by phosphorylating and activating PLC-Beta and Gamma. This process is also well regulated by BAM32. These Phospholipases utilize PIP2 as a substrate for the generation of IP3 (Inositol-1,4,5-trisphosphate) and DAG (Diacylglycerol). IP3 activates IP3R (IP3 Receptor) to release stored Calcium ions, whereas DAG and Calcium activate PKC-Beta (Protein Kinase-C-Beta) and Calm (Calmodulin), respectively. One of the targets regulated by calcium elevation is the transcription factor NFAT (Nuclear-Factor of Activated T-cells), whose nuclear translocation is facilitated through its dephosphorylation by Caln (Calcineurin) by Calm. PKC-Beta appears to phosphorylate IKKs (I-KappaB Kinases) activity resulting in the phosphorylation and degradation of I-KappaBs (Inhibitor of Kappa Light Chain Gene Enhancer in B-Cells) and translocation of NF-KappaB (Nuclear Factor-Kappa-B) to the nucleus. PKC activates BIMP1 (Bcl10-Interacting MAGUK Protein-1)/Bcl10 (B-Cell Cell/Lymphoma-10)/MALT1 (Mucosa Associated Lymphoid Tissue Lymphoma Translocation Gene-1) that forms a strong and specific complex within the cell to synergize in the activation of NF-KappaB, whereas, Calm activates CaMKII (Calcium/Calmodulin-Dependent Protein Kinase-II) to facilitate the same (Ref.9 & 10). Translocation of NF-KappaB by catalytic PI3Ks further enhances the BCR-stimulated PI3K gene expression. PI3K mediated gene expression under the influence of BCR/c-Abl activate the MAPK cascade via aPKC (Atypical PKC)/Raf1 (v-Raf1 Murine Leukemia Viral Oncogene Homolog-1)/MEK (MEK Kinase) route. Further activation of MAPKs regulate subsequent MAPK Pathways to control cell proliferation through induction of transcription factors (like NF-KappaB, NFAT, Elk1 (ETS-domain protein Elk1), ATFs (Activating Transcription Factors) and CREB (cAMP Response Element-Binding Protein)). These factors also regulate Opn (Osteopontin) gene expression that modulates cellular processes like immune homeostasis, ossification and myelination. Opn upon translocation to the exterior controls Integrin Pathway through ECM/Integrin interactions to recruit the regulatory PI3Ks (Ref.11).
The production of second messenger by PI3K and activation of intermediate signaling proteins is the concerted regulation of several transcription factors that mediate gene transcription in the B-cells. Since the immune response in vivo is regulated by alteration of these signaling outcomes, achieving a precise understanding of intracellular molecular events leading to B-lymphocyte proliferation, deletion, receptor editing, and survival still remains a challenge for the future. Understanding the contributions of PI3Ks to lymphocyte biology may contribute to the development of treatments for immune-related diseases, including autoimmunity, Leukaemia and graft rejection (Ref.12).
References
- 1
- Fruman DA. Phosphoinositide 3-kinase and its targets in B-cell and T-cell signaling.
- 2
- Okkenhaug K, Vanhaesebroeck B. PI3K in lymphocyte development, differentiation and activation.
- 3
- Donahue AC, Fruman DA. PI3K signaling controls cell fate at many points in B lymphocyte development and activation.
- 4
- Hebeis B, Vigorito E, Kovesdi D, Turner M. Vav proteins are required for B-lymphocyte responses to LPS.
- 5
- Otero DC, Omori SA, Rickert RC. Cd19-dependent activation of Akt kinase in B-lymphocytes.
- 6
- Tybulewicz VL. Commentary: New insights into the complexity of phosphatidylinositol lipid signaling in B lymphocytes.
- 7
- Decker SJ, Saltiel AR. Staying in SHIP shape.
- 8
- Marshall AJ, Krahn AK, Ma K, Duronio V, Hou S. TAPP1 and TAPP2 are targets of phosphatidylinositol 3-kinase signaling in B cells: sustained plasma membrane recruitment triggered by the B-cell antigen receptor.
- 9
- Hess KL, Donahue AC, Ng KL, Moore TI, Oak J, Fruman DA. Frontline: The p85alpha isoform of phosphoinositide 3-kinase is essential for a subset of B cell receptor-initiated signaling responses.
- 10
- Jou ST, Carpino N, Takahashi Y, Piekorz R, Chao JR, Carpino N, Wang D, Ihle JN. Essential, nonredundant role for the phosphoinositide 3-kinase p110delta in signaling by the B-cell receptor complex.
 关于我们
关于我们