NGF_Pathway
发布时间:2019-12-11 10:54 来源:SABiosciences
- 通路
- 概述
Review
One of the most fundamental issues in current biology is how to maintain the critical balance between cell survival and death, both during development and in adulthood. Unrestrained cell division and survival leads to various forms of tumor, while excessive or premature cell death may lead to a variety of diseases such as Alzheimer's, Parkinson's and Multiple Sclerosis. In the nervous system, a family of Neurotrophins, which includes NGF (Nerve Growth Factor), BDNF (Brain Derived Neurotrophic Factor), NT3 (Neurotrophin3) and NT4/5 (Neurotrophin-4/5), maintains this critical balance of cell survival and death. The best characterized of these is the NGF, a neurotrophic protein which is important for the growth and survival of diverse peripheral and central neurons. NGF is expressed within discrete CNS (Central Nervous System) areas and is secreted by tissues targeted by sympathetic and some sensory neurons. The effects of NGF on neuronal growth and survival are mediated by a high affinity cell surface receptor TRKA (Tyrosine Kinase Receptor-A) and a low affinity neurotrophic receptor, p75(NTR) (p75 Neurotrophic Receptor). NGF may signal its neuroprotective actions through the TRKA receptor and trigger apoptosis in some cells through the p75 receptor (Ref.1 & 2).
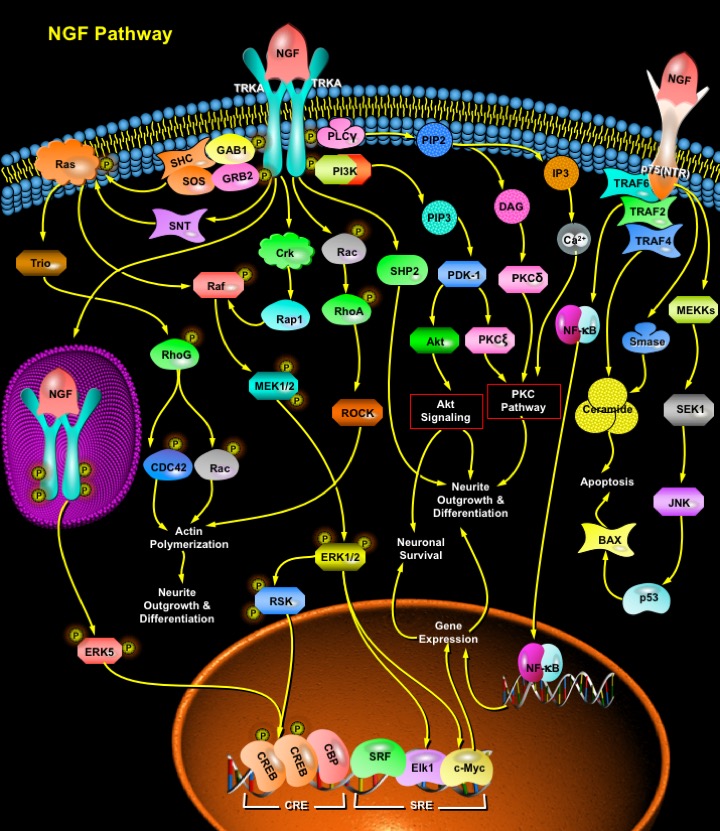
Activation of TRKA leads to a cascade of intracellular events with consequent changes in the expression of genes encoding diverse enzymes, ion channels and other cellular components. This activation of TRKA is followed by receptor dimerization and transautophosphorylation of tyrosine residues on TRKA, which act as docking sites for a variety of downstream signaling effectors and adaptor proteins. Activation of TRKA results in rapid association of TRKA with PLC-Gamma (Phospholipase-C-Gamma), PI3K (Phosphatidylinositiol-3 Kinase) and the adaptor proteins. TRKA tyrosine phosphorylates the adaptor proteins GAB1 (GRB2-Associated Binding Protein-1) and SHC (SH2 Containing Protein), resulting in their association with GRB2 (Growth Factor Receptor-Bound Protein-2)-SOS, a complex that enhances the rate of GDP-GTP exchange on Ras, leading to Ras activation. Activated Ras binds to Raf, which in turn, phosphorylates and activates MEK (MAPK/ERK Kinase). MEKs are dual specificity kinases which phosphorylate the MAPKs (Mitogen-Activated Proteins)/ ERK1/2 (Extracellular signal-regulated Kinases). Phosphorylated ERK1/2 then participate in at least two cascades. ERK1/2 may translocate into the nucleus, where they phosphorylate the transcription factor Elk1, or they phosphorylate the RSK (Ribosomal S6 Kinase). Phosphorylation of Elk1 allows it to interact with the accessory transcription factor SRF (Serum Response Factor), after which it binds to the SRE (Serum Response Element) within the c-Fos promoter region and contributes to the initiation of transcription. Phosphorylation of RSK leads to its nuclear translocation and consequent phosphorylation of CREB (Cyclic Adenosine Monophosphate (c-AMP) Response Element Binding Protein). Phosphorylated CREB binds to the transcriptional coactivator protein CBP (CREB Binding Protein) and the SRF-Elk complex creating an extended transcriptional factor complex leading to c-Fos transcription. The activation of transcription factors regulates the expression of NGF-inducible genes and contributes to NGF-induced neurite formation. Crk and SNT also modulate MAPK activity in some neuronal cells. The small G-protein Rap1 that lies downstream of Crk, links to the MAPK cascade by physically binding to and activating Raf, whereas, SNT contributes to Ras activation (Ref.2, 3, 4 & 5). Sometimes CREB phosphorylation requires activation and endocytosis of TRKAs located at the axon terminals. Within distal axons, activated TRKAs are internalized into vesicles. These vesicles give rise to signaling endosomes that are transported from distal axons back to the cell bodies. The binding of NGF to TRKA receptors at the axon terminal results in local activation of ERK1/2 and ERK5 within the axon terminal and also activation of ERK5 in the cell body. Components of the ERK5 cascade are transported in endosomes together with TRKAs. The activated ERK5 moves from the cell body to the nucleus, where it mediates CREB phosphorylation and gene expression. The ERK5 pathway is required for CREB phosphorylation following neurotrophin stimulation of distal axons, whereas both ERK1/2 and ERK5 contribute to CREB activation following neurotrophin stimulation of cell bodies (Ref.6).
After being recruited by TRKA, PLC-Gamma catalyses the formation of DAG (Diacylglycerol) and IP3 (Inositol Triphosphate) from PIP2 (Phosphatidylinositol 4,5-Bisphosphate). These second messengers: DAG and IP3 stimulate PKC-Delta (Protein Kinase-C-Delta) and increase intracellular Ca2+ (Calcium) respectively, which may play a role in cytoskeletal rearrangements thus contributing to neurite outgrowth and differentiation. Another signaling intermediate that forms a complex with activated TRKA is PI3K (Phosphatidylinositiol-3 Kinase). At the membrane PI3K converts PIP2 (Phosphatidylinositol 3,4-bisphosphate) to PIP3 (Phosphatidylinositol 3,4,5-Trisphosphate) which in turn, activates a protein kinase called PDK-1 (Phosphoinositol Dependent Kinase-1). PDK-1 phosphorylates and activates another enzyme Akt, that occupies the central position in the activation of cell survival pathways in many cells. The activation of Akt induces the activation of several downstream effectors that lead to survival and differentiation of neurons. PDK-1 also contributes to neurite outgrowth through the activation of PKC-Zeta (Protein Kinase-C-Zeta) (Ref.2 & 5). Rac, Rho and CDC42, members of the Rho family GTPase proteins, actively participate in neurite outgrowth and differentiation. Activated TRKA gives rise to Actin Polymerization, by activating the Rac-->RhoA-->ROCK (Rho-Associated Coiled-Coil-Containing Protein Kinase) pathway. Actin polymerization and formation of filopodia or lamellipodia are important events leading to neurite differentiation. Another type of Rho family GTPase, RhoG, transduces the Ras signal to Rac1 and Cdc42 activation in neuronal cells. Trio (Triple Functional Domain-PTPRF Interacting) is an upstream regulator of RhoG for the NGF-induced neurite outgrowth. Trio-->RhoG-->Rac1 and Trio-->RhoG-->Cdc42 pathways form a central signaling route for neurite outgrowth (Ref.7).
NGF also binds, with lower affinity relative to TrkA, to the p75(NTR). p75(NTR) is a member of the TNFR (Tumor Necrosis Factor) receptor superfamily, having no tyrosine kinase domain. p75(NTR) activates a survival pathway that may involve the activation of NF-KappaB or the activation of the JNK (Jun N-terminal Kinase)/c-Jun death pathway. The main physiological functions of p75NTR are to activate TRKA-independent signal transduction cascades involving Sphingomyelinase, NF-KappaB (Nuclear Factor-KappaB, and JNK. p75NTR activates a survival pathway that may involve the activation of NF-KappaB through TRAF6 (TNF Receptor Associated Factor-6), whereas in limiting amounts of NGF p75NTR may activate the JNK death pathway. Regulation of NF-KappaB-dependent transcription occurs mainly via cytosolic retention by a family of inhibitory molecules known as I-KappaB. Degradation of I-KappaB releases NF-KappaB molecules that in turn translocate into the nucleus to evoke their response. NF-KappaB transcription factors are a family of dimeric molecules, including NF-KappaB1 (p50/p105), NF-KappaB2 (p52/p100), RelA, RelB, and c-Rel, all of which are involved in regulating cell survival. NGF binding selectively to p75(NTR) brings about the activation of the proapoptotic JNK cascade. MEKKs (MAP/ERK Kinase Kinases) and SEK ((SAPK/ERK Kinase) are the upstream regulators of JNK. JNKs, in turn, upregulate p53 and the proapoptotic member of the Bcl2 family: BAX in a sequencial manner, which bring about apoptosis of the neuronal cells. p75(NTR) activation by NGF also recruits several TRAFs (TRAF2 and TRAF4) to the receptor that are involved in Ceramide generation. Ceramide is a key player in the induction of neuronal apoptosis. It can also be generated by p75(NTR) by the activation of Sphingomyelinase (Ref.8, 9, 10 & 11).
NGF is the most potent growth factor for cholinergic neurons and it influences the proliferation, differentiation, survival and death of neuronal cells. It is essential for the health and well-being of the nervous system and is a promising candidate for treating Alzheimer’s disease. Alterations in NGF levels have been implicated in neurodegenerative disorders, such as Alzheimer's disease and Huntington's disease, as well as psychiatric disorders, including depression and substance abuse. Signals emanating from receptors for NGF control practically all aspects of immune defense and, as such, constitute potential targets for therapeutic intervention through rational drug design in neurodegenerative conditions (Ref.3, 9 & 12).
References
- 1
- Sofroniew MV, Howe CL, Mobley WC. Nerve growth factor signaling, neuroprotection, and neural repair.
- 2
- Freund-Michel V, Bertrand C, Frossard N. TrkA signalling pathways in human airway smooth muscle cell proliferation.
- 3
- Chao MV, Rajagopal R, Lee FS. Neurotrophin signalling in health and disease.
- 4
- Cheung EC, Slack RS. Emerging role for ERK as a key regulator of neuronal apoptosis.
- 5
- Schramm A, Schulte JH, Astrahantseff K, Apostolov O, Limpt V, Sieverts H, Kuhfittig-Kulle S, Pfeiffer P, Versteeg R, Eggert A. Biological effects of TrkA and TrkB receptor signaling in neuroblastoma.
- 6
- Watson FL, Heerssen HM, Bhattacharyya A, Klesse L, Lin MZ, Segal RA. Neurotrophins use the Erk5 pathway to mediate a retrograde survival response.
- 7
- Negishi M, Katoh H. Rho family GTPases as key regulators for neuronal network formation.
- 8
- Schor NF. The p75 neurotrophin receptor in human development and disease.
- 9
- Wallach D, Arumugam TU, Boldin MP, Cantarella G, Ganesh KA, Goltsev Y, Goncharov TM, Kovalenko AV, Rajput A, Varfolomeev EE, Zhang S. How are the regulators regulated? The search for mechanisms that impose specificity on induction of cell death and N
- 10
- Yuan J, Yankner BA. Apoptosis in the nervous system.
 关于我们
关于我们