JAK_STAT_Pathway
发布时间:2019-12-11 09:17 来源:SABiosciences
- 通路
- 概述
Review
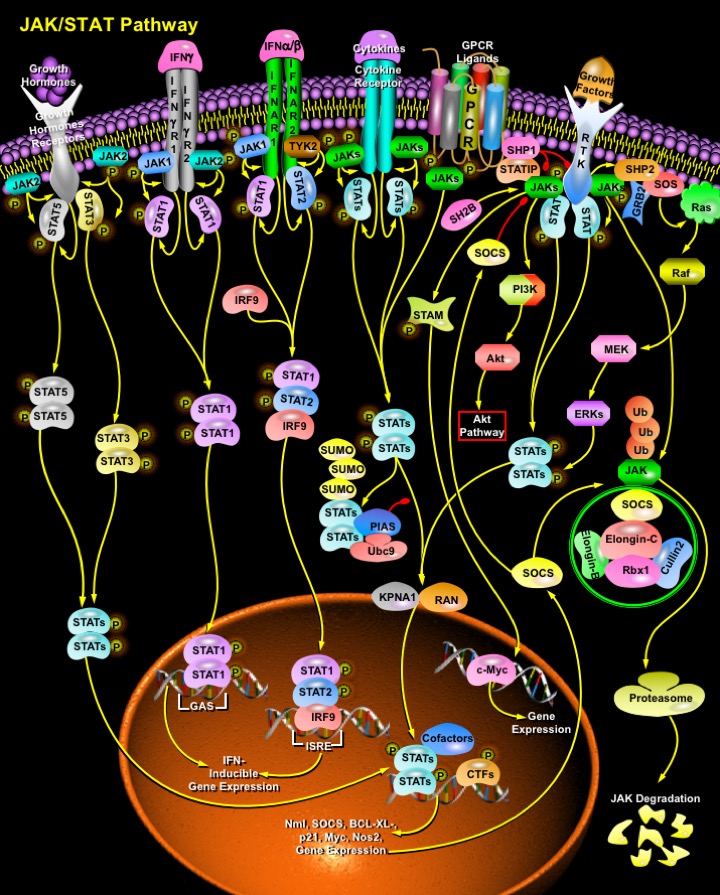
Signaling pathways mediating the transduction of information between cells are essential for development, cellular differentiation and homeostasis. Their dysregulation is also frequently associated with human malignancies. The JAK (Janus tyrosine Kinase)-STAT (Signal Transducer and Activator of Transcription) pathway represents one such signaling cascade whose evolutionarily conserved roles include cell proliferation and haematopoiesis. JAK belongs to a family of non-receptor protein tyrosine kinases of approximately 130 kDa, comprising of JAK1, JAK2, JAK3 and TYK2 (non-receptor Protein Tyrosine Kinase-2). STATs are latent cytoplasmic transcription factors that become activated after recruitment to an activated receptor complex. Seven STAT proteins have been identified, STAT1 to 6, including STAT5a and STAT5b, which are encoded by distinct genes. In addition, different isoforms of several STATs have been identified. Evolutionarily conserved in eukaryotic organisms from slime molds to humans, JAK-STAT signaling appears to be an early adaptation to facilitate intercellular communication that has co-evolved with myriad cellular signaling events. This co-evolution has given rise to highly adapted, ligand-specific signaling pathways that control gene expression. In addition, the JAK-STAT signaling pathways are regulated by a vast array of intrinsic and environmental stimuli, which can add plasticity to the response of a cell or tissue (Ref.1 & 2).
Mechanistically, JAK/STAT signaling is relatively simple, with only a few principal components. A variety of ligands including Cytokines, Hormones and Growth factors, and their receptors stimulate the JAK/STAT pathway. Intracellular activation occurs when ligand binding induces the multimerization of receptor subunits. For some ligands, such as Epo (Erythropoietin) and GH (Growth Hormone), the receptor subunits are bound as homodimers while, for others, such as Ifns (Interferons) and ILs (Interleukins), the receptor subunits are heteromultimers. For signal propagation, the cytoplasmic domains of two receptor subunits must be associated with JAK tyrosine kinases. JAK activation occurs upon ligand-mediated receptor multimerization because two JAKs are brought into close proximity, allowing trans-phosphorylation. The activated JAKs subsequently phosphorylate additional targets, including both the receptors and the major substrates, STATs. The seven mammalian STATs bear a conserved tyrosine residue near the C-terminus that is phosphorylated by JAKs. This phosphotyrosine permits the dimerization of STATs through interaction with a conserved SH2 domain. Different JAKs and STATs are activated by different ligands. For example, Hormones such as GH, Epo and Tpo (Thrombopoietin) generally stimulates the activation of JAK2 as well as STAT3 and 5. Phosphorylated STATs then enter the nucleus by a mechanism that is dependent on Importin Alpha-5 (also called nucleoprotein interactor 1) and the Ran nuclear import pathway. Once in the nucleus, dimerized STATs bind specific regulatory sequences to activate or repress transcription of target genes. Thus the JAK/STAT cascade provides a direct mechanism to translate an extracellular signal into a transcriptional response. RTKs (Receptor Tyrosine kinases) commonly activate Ras/Raf/MEK/ERK signaling but when overactivated can also induce the JAK/STAT pathway, originally identified as the signaling cascade downstream of cytokine receptors (Ref.3 & 4).
Cytokines and their receptors are the major activator of JAK/STAT pathway. Ifns are antiviral cytokines that are produced by many cell types following viral infection. Ifns are classified as either Type I or Type II. Type I Ifns include Ifn-Alpha, -Beta, -Omega and -Tau, which are all monomeric; the only Type II Ifn is Ifn-Gamma, a dimer. Type I and II Ifns bind to different receptors and activate overlapping but distinct genes. Ifn are very important regulator of JAK/STAT pathway. The Ifn-AlphaBetaR consists of two subunits, IfnAR1 and IfnAR2, which form a heterodimer upon Ifn stimulation. This initiates the activation of two Janus-family tyrosine kinases; JAK1 and TYK2 followed by phosphorylation of STAT1 and STAT2 (Signal Transducers and Activators of Transcription) proteins. The phosphorylated STATs then dissociate from the receptor heterodimer and bind to IRF9/p48, a member of the IRF (Interferon Regulatory Factor-9) family, forming a trimeric major interferon gene factor known as ISGF3 (Interferon-Stimulated Transcription Factor-3) complex. This complex translocates to the nucleus and binds to the cis element ISRE (Ifn-stimulated Response Element) thereby initiating transcription of several Ifn-inducible genes. In contrast, binding of Ifn-Gamma to its receptor leads to tyrosine phosphorylation of the JAK1 and JAK2 tyrosine kinases, resulting in the phosphorylation of STAT1 but not STAT2. Phosphorylated STAT1 homodimerizes to form the GAF-AAF complex, which translocates to the nucleus and binds to the GAS element present in most Ifn-Gamma inducible genes (Ref.5 & 6).
In addition to the principal components of the pathway, other effector proteins have been identified that contribute to at least a subset of JAK/STAT signaling events. STAMs (signal-transducing adapter molecules) are adapter molecules with conserved VHS and SH3 domains. STAM1 and STAM2A can be phosphorylated by JAK1-JAK3 in a manner that is dependent on a third domain present in some STAMs, the ITAM (inducible tyrosine-based activation motif). Through a poorly understood mechanism, the STAMs facilitate the transcriptional activation of specific target genes, including Myc. A second adapter that facilitates JAK/STAT pathway activation is STATIP (STAT-Interacting Protein), a WD40 protein. STATIPs can associate with both JAKs and unphosphorylated STATs, perhaps serving as a scaffold to facilitate the phosphorylation of STATs by JAKs. A third class of adapter with function in JAK/STAT signaling is the SH2B/Lnk/APS family. These proteins contain both pleckstrin homology and SH2 domains and are also substrates for JAK phosphorylation. Both SH2-Bß and APS associate with JAKs, but the former facilitates JAK/STAT signaling while the latter inhibits it (Ref.1 & 7).
In addition to JAK/STAT pathway effectors, there are three major classes of negative regulator: SOCS (suppressors of cytokine signaling), PIAS (protein inhibitors of activated STATs) and PTPs (protein tyrosine phosphatases). Perhaps the simplest are the tyrosine phosphatases, which reverse the activity of the JAKs. The best characterized of these is SHP-1. SHP-1 contains two SH2 domains and can bind to either phosphorylated JAKs or phosphorylated receptors to facilitate dephosphorylation of these activated signaling molecules. Other tyrosine phosphatases, such as CD45, appear to have a role in regulating JAK/STAT signaling through a subset of receptors. The second class of negative regulators includes SOCS. SOCS proteins are a family of at least eight members containing an SH2 domain and a SOCS box at the C-terminus. The hallmark of the SOCS family is the SOCS Box, which mediates interaction with the Elongin-B/C complex and couples the SOCS and associated target proteins JAKs to the proteasomal protein degradation pathway. The NH2-terminus of the SOCS Box contains a conserved Elongin-B/C binding motif (BC Box) that binds to Elongin-C, which in turn associates with a complex consisting of Elongin-B, a Cullin family member, and the RING finger protein Rbx-1 to form a multiprotein complex capable of acting as an E3 ubiquitin ligase. Together with an ATP-dependent ubiquitin-activating enzyme (E1) and an ubiquitin-conjugating enzyme (E2), the E3 ubiquitin ligase acts to tag proximal proteins with polyubiquitin chains. Polyubiquitination targets proteins for degradation by the proteasome. Ubiquitin is then recycled by dUB (deubiquitinating enzymes) and the cycle continues. The SOCS complete a simple negative feedback loop in the JAK/STAT circuitry: activated STATs stimulate transcription of the SOCS genes and the resulting SOCS proteins bind phosphorylated JAKs and their receptors to turn off the pathway. The third class of negative regulator is the PIAS proteins: PIAS1, PIAS3, PIASx and PIASy. The PIAS proteins bind to activated STAT dimers and prevent them from binding DNA. The mechanism by which PIAS proteins act remains unclear. However, PIAS proteins have recently been demonstrated to associate with the E2 conjugase Ubc9 and to have E3 conjugase activity for sumoylation that is mediated by the RING finger domain. Although there is evidence that STATs can be modified by sumoylation, the function of that modification in negative regulation is not yet known (Ref.8, 9 & 10).
Although the mechanism of JAK/STAT signaling is relatively simple, the biological consequences of pathway activation are complicated by interactions with other signaling pathways. JAK/STAT signaling also indirectly promotes Ras signaling through the transcriptional activation of SOCS3. SOCS3 binds RasGAP, a negative regulator of Ras signaling, and reduces its activity, thereby promoting activation of the Ras pathway. Reciprocally, RTK pathway activity promotes JAK/STAT signaling by at least two mechanisms. First, the activation of some RTKs, including EGFR and PDGFR, results in the JAK-independent tyrosine phosphorylation of STATs, probably by the Src kinase. Second, RTK/Ras pathway stimulation causes the downstream activation of MAPK. MAPK specifically phosphorylates a serine near the C-terminus of most STATs. Furthermore, the functions of activated STATs can be altered through association with other transcription factors (c-Jun,IRF9 (Interferon Regulatory Factor-9), c-Fos, NF-KappaB (Nuclear Factor-KappaB), SMAD(Sma and MAD (Mothers Against Decapentaplegic) Related Protein), SP1 (Transcription Factor SP1)) and cofactors like p300, CBP (CREB-Binding Protein), BRCA1 (Breast Cancer-1 Gene), MCM5, (Minichromosome Maintenance-5), that are regulated by other signaling pathways. In addition to activating STATs, JAK kinases phosphorylate other signaling/adaptor proteins, linking JAK signaling to other pathways such as the MAP Kinases. JAKs or STATs can also participate in signaling through other receptor classes. STATs are also involved in many diseases like breast cancer (tumors), head and neck cancer (cell lines), multiple myeloma, Erythro leukemia, acute myelocytic leukemia, EBV-related Burkitt’s lymphoma, and herpes virus salmiri–dependent lymphoma (Ref.11 & 12).
References
- 1
- Ananthakrishnan R, Hallam K, Li Q, Ramasamy R. JAK-STAT pathway in cardiac ischemic stress.
- 2
- Walker JG, Smith MD. The Jak-STAT pathway in rheumatoid arthritis.
- 3
- Marrero MB. Introduction to JAK/STAT signaling and the vasculature.
- 4
- Gao B. Cytokines, STATs and liver disease.
- 5
- Rani MR, Ransohoff RM. Alternative and accessory pathways in the regulation of IFN-beta-mediated gene expression.
- 6
- Best SM, Morris KL, Shannon JG, Robertson SJ, Mitzel DN, Park GS, Boer E, Wolfinbarger JB, Bloom ME. Inhibition of interferon-stimulated JAK-STAT signaling by a tick-borne flavivirus and identification of NS5 as an interferon antagonist.
- 7
- Valentino L, Pierre J. JAK/STAT signal transduction: regulators and implication in hematological malignancies.
- 8
- Espert L, Dusanter-Fourt I, Chelbi-Alix MK. Negative regulation of the JAK/STAT: pathway implication in tumorigenesis.
- 9
- Weniger MA, Melzner I, Menz CK, Wegener S, Bucur AJ, Dorsch K, Mattfeldt T, Barth TF, Moller P. Mutations of the tumor suppressor gene SOCS-1 in classical Hodgkin lymphoma are frequent and associated with nuclear phospho-STAT5 accumulation.
- 10
- Rakesh K, Agrawal DK. Controlling cytokine signaling by constitutive inhibitors.
 关于我们
关于我们