IL-10
发布时间:2019-12-10 16:10 来源:SABiosciences
- 通路
- 概述
Review
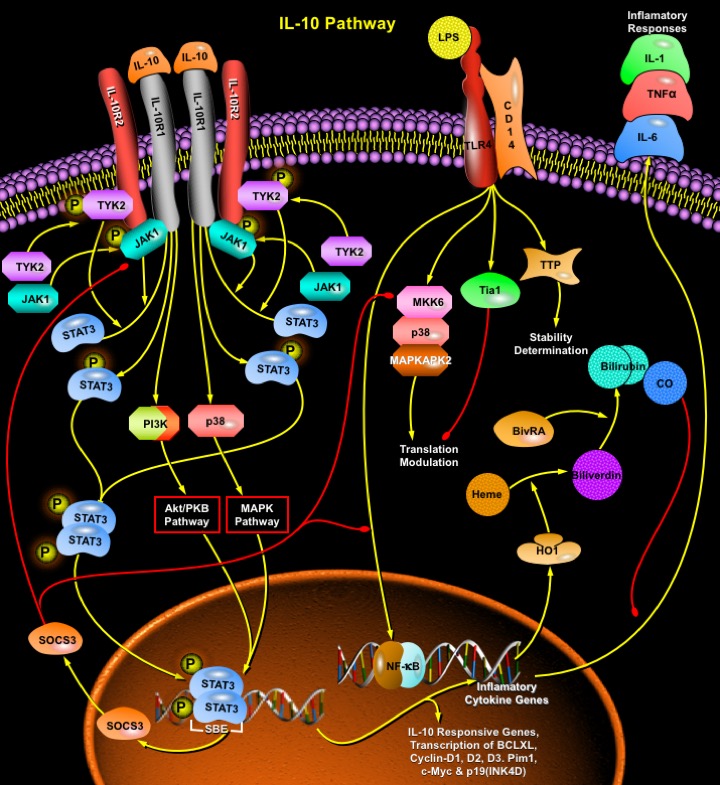
IL-10 (Interleukin-10) is a pleiotropic cytokine with important immunoregulatory functions whose actions influence activities of many of the cell-types in the immune system. It is a cytokine with potent anti-inflammatory properties, repressing the expression of inflammatory cytokines such as TNF-Alpha (Tumor Necrosis Factor-Alpha), IL-6 (Interleukin-6) and IL-1 (Interleukin-1) by activated macrophages. Functional IL-10R (IL-10 Receptor) complexes are tetramers consisting of two ligand-binding subunits (IL-10R-Alpha or IL-10R1) and two accessory signaling subunits (IL-10R-Beta or IL-10R2). Binding of IL-10 to the extracellular domain of IL-10R1 activates phosphorylation of the receptor-associated, JAK1 (Janus Kinase-1) and TYK2 (Tyrosine Kinase-2), which are constitutively associated with IL-10R1 and IL-10R2, respectively. These kinases then phosphorylate specific tyrosine residues (Y446 and Y496) on the intracellular domain of the IL-10R1 chain. Once phosphorylated, these tyrosine residues (and their flanking peptide sequences) serve as temporary docking sites for the latent transcription factor, STAT3 (Signal Transducer and Activator of Transcription-3). STAT3 binds to these sites via its SH2 (Src Homology-2) domain, and is, in turn, tyrosine-phosphorylated by the receptor-associated JAKs. It then homodimerizes and translocates to the nucleus where it binds with high affinity to SBE (STAT-Binding Elements) in the promoters of various IL-10-responsive genes. Constitutively active forms of STAT3 increase transcription of anti-apoptotic and cell-cycle-progression genes such as BCLXL, Cyclin-D1, Cyclin-D2, Cyclin-D3, and Cyclin-A, Pim1, c-Myc and p19(INK4D) (Ref.1).
IL-10 has also been reported to activate another major survival pathway consisting of PI3K (Phosphoinositide-3 Kinase) and its downstream substrates p70S6K (p70 S6-kinase) and Akt/PKB (Protein Kinase-B). Although the anti-inflammatory effects of IL-10 are not mediated via PI3K, the ability of IL-10 to promote survival of astrocytes or to induce proliferation of mast cells depends upon the activation of PI3K (Ref.2).
IL-10 also interferes with the activation potential of the p38/MAPK (Mitogen-Activated Protein Kinase) pathway, which is required to activate TNF (Tumor Necrosis Factor) translation. IL-10-mediated signals primarily target TNF-mRNA translation. This process is clearly dependent on TNF-3'ARE. IL-10 leads to ARE-dependent destabilization of granulocyte–macrophage colony-stimulating factor, granulocyte colony-stimulating factor and KC cytokine mRNAs. Inhibition of p38/MAPK affects TNF-mRNA translation by inhibiting LPS (Lipopolysaccharide)-induced polysome coupling of TNF-mRNA in macrophages, without affecting TNF-mRNA accumulation. Similarly, the absence of MK2 leads to a profound reduction of LPS-induced TNF production, yet TNF-mRNA accumulation and stability are not affected. Thirdly, and most importantly, TTP (TNF-ARE Binding Protein Regulating-TNF Biosynthesis) exerts a potent destabilizing activity on TNF-mRNA without imposing translational control. Two RNA binding proteins that have recently been demonstrated to have TNF-ARE binding capacities are TiaR and Tia1 (Ref.3). The detailed mechanism by which IL-10 targets MAPK/SAPK (Stress-Activated Protein Kinase) signals is currently unknown. It is possible that IL-10 signaling interferes directly with the activation of MAPKs. There is evidence that points towards the requirement for JAK1/STAT3 in TNF suppression. Thus, one possibility would be that STAT3 activates factors capable of interfering with MAPK/SAPK signaling. One of the IL-10-responsive genes, SOCS3 (Suppressor of Cytokine Signaling-3) is a member of a newly identified family of genes that inhibit JAK/STAT-dependent signaling. Moreover, the ability of IL-10 to induce de novo synthesis of SOCS3 in monocytes correlates with its ability to inhibit expression of many genes in these cells, including endotoxin-inducible cytokines such as TNF-Alpha and IL-1. Both STAT3 and SOCS3 can be activated by inflammatory stimuli like LPS and TNF and, most importantly, SOCS3 activation can also be induced by STAT3-independent mechanisms. In addition, LPS can synergize with IL-10 to prolong SOCS3 mRNA stability in myeloid cells. Therefore, early post-transcriptional mechanisms may provide a pool of SOCS3, which can be rapidly activated in the absence of de novo gene transcription. Consequently, SOCS3 transcription may allow for the replenishment of a SOCS3 pool to enforce its negative action in a temporal manner, thus showing p38/MAPK dependency of both STAT3 and SOCS3 for their activation as well as for the apparent capacity of SOCS3 to suppress STAT3 activation itself (Ref.4).
IL-10 is a potent inducer of HO1 (Heme Oxygenase-1) in murine primary macrophages and J774 cell line. The induction of HO1 occurs at transcriptional level, and is mediated via a p38 MAP-kinase–dependent pathway. HO1 is involved in the biosynthesis of heme, and catalyzes a reaction producing CO (Carbon-monoxide), free iron, and the heme precursor biliverdin. Inhibition of HO1 protein synthesis by antisense oligonucleotide significantly reversed the inhibitory effect of IL-10 on the production of TNF-Alpha induced by LPS. HO1 induction is also essential for the suppressive effect of IL-10 on LPS-induced expression of inducible nitric oxide synthase as well as matrix metalloproteinase-9. CO derived from heme degradation by HO1 mediates the anti-inflammatory effect of IL-10 in macrophages. IL-10 also promotes growth and survival of cancer cells, including non-Hodgkin's lymphoma, Burkitt lymphoma and non-small cell lung cancer. Recently, IL-10 has been demonstrated to directly increase survival of both cortical and cerebellar granule neurons, astrocytes and progenitor as well as differentiated oligodendrocytes (Ref.5).
References
- 1
- Donnelly RP, Dickensheets H, Finbloom DS. The interleukin-10 signal transduction pathway and regulation of gene expression in mononuclear phagocytes.
- 2
- Strle K, Zhou JH, Broussard SR, Venters HD, Johnson RW, Freund GG, Dantzer R, Kelley KW. IL-10 promotes survival of microglia without activating Akt.
- 3
- Kontoyiannis D, Kotlyarov A, Carballo E, Alexopoulou L, Blackshear PJ, Gaestel M, Davis R, Flavell R, Kollias G. Interleukin-10 targets p38 MAPK to modulate ARE-dependent TNF mRNA translation and limit intestinal pathology.
- 4
- Meisel C, Vogt K, Platzer C, Randow F, Liebenthal C, Volk HD. Differential regulation of monocytic tumor necrosis factor-alpha and interleukin-10 expression.
- 5
- Tzong-Shyuan Lee and Lee-Young Chau. Heme oxygenase-1 mediates the anti-inflammatory effect of interleukin-10 in mice.
 关于我们
关于我们