Chemokine_Signaling
发布时间:2019-12-09 19:58 来源:SABiosciences
- 通路
- 概述
Review
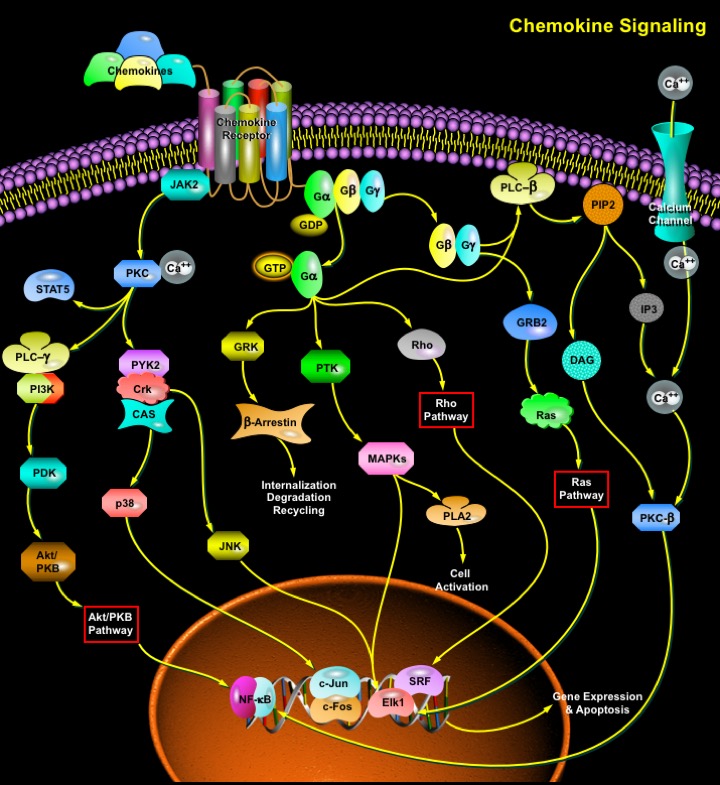
Chemokines, or chemotactic cytokines, are a large family of small (6–14 kDa), structurally related proteins that mediate a wide range of biological activities. As a part of normal immune system functions, chemokines are a critical component of basal leukocyte trafficking essential for immune system architecture and development, and immune surveillance. Chemokines also participate in the growth, differentiation, and activation of leukocytes as well as stimulate various effector functions of these cells, such as integrin activation, chemotaxis, superoxide radical production and granule enzyme release. Four classes of chemokines have been defined by the arrangement of the conserved cysteine (C) residues of the mature proteins: the CXC chemokines the CC chemokines in which the first two conserved cysteines residues are adjacent; the C chemokines that lack two (the first and third) of the four conserved cysteine residues; and the CX3C chemokines which have three intervening AA residues between the first two conserved cysteine residues.
Chemokines mediate their effects through interactions with seven-transmembrane-spanning glyco-protein receptors coupled to a G-protein signaling pathway. Chemokine receptors are promiscuous, as each can bind more than one chemokine; expression is heterogeneous among different cells of the leukocyte lineage and is transcriptionally regulated (Ref.1). G-proteins are inactive when GDP is bound to the G-protein subunit, but they become active when GDP is exchanged for GTP. During ligand binding, chemokine receptors associate with G-proteins, facilitating the exchange of GDP (Guanosine Diphosphate) for GTP (Guanosine Triphosphate). In the active state, G-proteins dissociate into G-alpha and G-beta gamma subunits, which are able to activate signaling molecules that mediate changes in the cytoskeletal apparatus, and transcription factors that regulate cell growth. Activated beta gamma subunits of G-Proteins trigger a series of reactions, commencing with the hydrolysis of PIP2 (Phosphatidylinositol-4,5-Bisphosphate) generating IP3 (1,4,5-Inositol Trisphosphate) and DAG (1,2-Diacylglycerol), an immediate consequence of which is the release of calcium from intracellular stores. In leukocytes, the signalling is mediated by Gi subunits, and the beta gamma subunits activate the PLC-Beta2 (Phospholipase-C-Beta2) enzyme to hydrolyse PIP2. This induction of calcium mobilization is one of the activities very often used to determine chemokine activity. There is increasing evidence that chemokine receptors can also activate several different intracellular effectors downstream of G-alphaI coupling, including the low-molecular-weight proteins Ras and Rho, PLA2 (Phospholipase-A2), PI3K (Phosphatidylinositol-3 Kinase), tyrosine kinases and the MAPK (Mitogen Activated Protein Kinase) pathway (Ref.2).
After binding to their specific receptors, chemokines may also induce receptor homodimerization and subsequently activate the receptor-associated JAK (Janus Kinases), possibly by transphosphorylation on tyrosine residues. This may create SH2 docking sites, leading to the recruitment of STAT (Signal Transducers and Activators of Transcription Factors) (Ref.3). Another group of GPCR (G Protein-Coupled Receptor), the opioid receptors, were recently shown to undergo heterodimerization. In this case, there is clear biochemical and pharmacological evidence for the heterodimerization of two functional opioid receptors, k and d. Heterodimerization of these receptors causes synergistic agonist binding and potentiates the biological signal (Ref.4).
Recently, chemokine receptors have also been identified as an essential component for the infection of host cells by HIV (Human Immunodeficiency Virus). Chemokine receptors thus present an attractive therapeutic target for inflammatory disorders and HIV infection. Chemokine receptors are essential for triggering chemotaxis to immune cells; however, a number of them can also mediate death when engaged by nonchemokine ligands. When the chemokine receptor CXCR4 is engaged by SDF1 (Stromal Cell-Derived Factor1) alpha, it triggers cells to chemotax, and in some cell types such as neurons, causes cell death. CXCR4 activation in CD4 T-Cells by SDF1-alpha led to the activation of the prosurvival second messengers, Akt and ERKs (Extracellular Signal Regulated Kinases). Chemokine biology lies at the critical interface between normal immune function and immune related disease. The large number of chemokines and receptors, the complex nature of their distinct and overlapping binding specificities, and the highly regulated expression of these proteins provide the opportunity to develop new types of therapeutics that selectively target subsets of leukocytes for the treatment of a variety of pathophysiological processes including chronic and acute inflammation, infectious diseases, and modulation of angiogenesis and fibrosis which are at the core of diseases as diverse as rheumatoid arthritis, atherosclerosis, multiple sclerosis, and AIDS (Ref.5).
References
- 1
- Amanda E.I. PROUDFOOT. The chemokine family. Potential therapeutic targets from allergy to HIV infection.
- 2
- Murdoch C, Finn A. Chemokine receptors and their role in inflammation and infectious diseases.
- 3
- Mellado M, Rodriguez-Frade JM, Vila-Coro AJ, de Ana AM, Martinez-A C. Chemokine control of HIV-1 infection.
- 4
- Mellado M, Rodriguez-Frade JM, Vila-Coro AJ, Fernandez S, Martin de Ana A, Jones DR, Toran JL, Martinez-A C. Chemokine receptor homo- or heterodimerization activates distinct signaling pathways.
- 5
- Vlahakis SR, Villasis-Keever A, Gomez T, Vanegas M, Vlahakis N, Paya CV. G protein-coupled chemokine receptors induce both survival and apoptotic signaling pathways.
 关于我们
关于我们